
Come sempre accade nel periodo primaverile-estivo, con l’allungarsi del fotoperiodo, che influisce positivamente sulla produzione di IGF-1 e di prolattina e negativamente su quella di melatonina da parte del sistema neuroendocrino delle bovine, si riscontra un aumento, a parità di lunghezza di giorni di lattazione, della produzione di latte concomitante ad una diminuzione dapprima del titolo di grasso e quindi di quello di proteina nel latte.
Mentre il secondo si può in buona parte attribuire al calo dell’ingestione di sostanza secca che avviene nei mesi più caldi, con una diminuzione della produzione di proteina batterica giornaliera legata al minor quantitativo di sostanza organica fermentata, e quindi necessita di interventi molto sottili sulla formulazione della razione, (apporto di amminoacidi ruminoprotetti, aumento della quota di RUP nella razione), per il titolo di grasso, che è l’argomento di questo articolo, la strategia volta a controbilanciare l’abbassamento del grasso stesso può essere articolata su più fronti.
Sappiamo infatti che la composizione acidica del grasso nel latte di una bovina è costituita da 3 macro frazioni:
- Gli acidi grassi de novo, ossia quelli sintetizzati nella mammella a partire dall’acido acetico prodotto nel rumine per allungamento della catena carboniosa grazie al acetilCoA carbossilasi e l’acido grasso sintetasi, oltre ad una piccola quota di acido butirrico che entra in circolo direttamente dalla parete ruminale. Questi acidi grassi sono a catena corta, fino a 15 atomi di carbonio, e i più rappresentati sono l’acido laurico (C12:0) e l’acido miristico (C14:0). Si considera normale una percentuale dal 21 al 25% del totale dei grassi.
- Gli acidi grassi misti, così detti perché in parte provenienti dall’elongazione dell’acido acetico in mammella e in parte derivanti direttamente dal tubo digerente, essenzialmente l’acido palmitico e i suoi isomeri. Una percentuale fisiologica di questi grassi sul totale del grasso nel latte è dal 35 al 41% del totale.
- Infine gli acidi grassi a 18 atomi di carbonio (preformati) derivanti dall’ingestione, che subiscono a livello ruminale un processo completo od incompleto di idrogenazione degli acidi grassi stessi oppure provenienti dalla mobilizzazione degli acidi grassi corporei. Questo gruppo è il più facilmente variabile, dipendendo dal contenuto in grassi totale della razione e dallo stato fisiologico (molto più elevati nel postparto) e va dal 30 al 45%.
Tramite un’analisi delle frazioni del grasso nel latte si può operare nei confronti della frazione carente (de novo e misti) o agire sul processo d’idrogenazione degli acidi grassi preformati e/o limitarne la mobilizzazione dai grassi di deposito.
Un contenuto elevato di grassi de novo, essendo indice di una buona efficienza di fermentazione, è correlato positivamente con il tasso di proteine nel latte dal momento che si ipotizza un’altrettanto elevata efficienza di produzione di biomassa microbic; inoltre, una quantità adeguata di de novo è correlata ad un’elevata frequenza di distribuzione del TMR. Naturalmente nell’immediato post-partum i grassi de novo nel latte sono molto bassi in quanto l’ingestione della bovina è bassa e il contenuto di NEFA nel sangue è molto alto.
Per quanto riguarda il contenuto di acido palmitico nel latte, esso può essere prodotto a partire dal l’acido acetico o tramite assunzione alimentare.
Nella dieta delle bovine gli acidi grassi a 18 atomi di carbonio vengono assunti in forma mono (MUFA) o poli (PUFA) insatura.
Per prevenire le carenze delle singole frazioni di acidi grassi ruminali si possono utilizzare diverse strategie:
1 – Acidi grassi de novo: favorire l’ingestione di sostanza secca, garantire un rapporto coretto tra acido acetico, propionico e butirrico a livello ruminale utilizzando razioni con fibra effettiva adeguata ottenuta tramite una maggior lunghezza di taglio. Questa procedura permette inoltre di avere meno grassi liberi nel caso dei foraggi insilati. L’utilizzo di foraggi con digeribilità della fibra elevata e costante nel corso della loro permanenza ruminale tende a migliorare ulteriormente questo aspetto.
Un altro aspetto interessante è il contenuto di uNDF, ossia la frazione fibrosa indigerita, che permette di mantenere un buon riempimento ruminale ed ha effetti meccanici interessanti. E’ stato di recente coniato un parametro di valutazione che tiene conto di questi due parametri ossia la PEuNDF.
PEuNDF = % PENDF x uNDF% PV
PEuNDF | uNDF% PV = 0.30 | uNDF% PV = 0.33 | uNDF% PV = 0.366 |
|---|---|---|---|
| % PENDF = 0.22 | 0.066 | 0.072 | 0.080 |
| % PENDF = 0.20 | 0.060 | 0.066 | 0.073 |
| % PENDF = 0.18 | 0.054 | 0.059 | 0.066 |
Come si vede dalla tabella, il valore corretto è attorno al 6,5% e può essere ottenuto aumentando la lunghezza di taglio di foraggi con basso uNDF, o accorciandola in caso di foraggi con uNDF alta.
L’utilizzo di sostanze che garantiscono anaerobiosi ruminale (lieviti vivi) o che aiutino a rompere il legame ligno cellulosico (aspergillus oryzae) può essere in certi casi di grande aiuto.
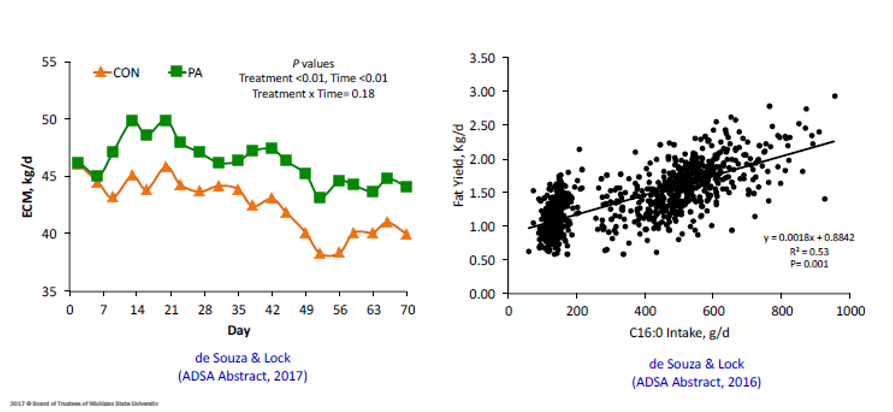
2 – Acido palmitico: il caso dell’acido palmitico è sicuramente il più semplice, dal momento che in caso di carenza si può intervenire con un maggior apporto alimentare dal momento che, essendo saturo, questo acido grasso attraversa il rumine senza variazioni e si rende disponibile come tale per l’assorbimento intestinale. Qui sotto sono riassunti gli effetti dell’ acido palmitico sulla produzione di grasso.
Figura 1 – Effetto dell’assunzione di C16:0 sulla produzione di latte corretta per il grasso e per l’energia.
3 – Più complicata e sottile è la strategia da adottare nei confronti della frazione di C:18 del latte.
I MUFA e i PUFA nel rumine vengono progressivamente saturati cambiando contestualmente la conformazione fisica dei doppi legami: tipicamente l’acido linoleico (C18:2), il più abbondante nella dieta delle bovine moderne, viene prima trasformato in acido rumenico cambiando la conformazione di un doppio legame da cis a trans, e successivamente in acido vaccenico (C18:1) ed infine ad acido stearico (C18:0). Più semplice è il destino dell’acido oleico (C18:1), che viene trasformato in stearico in una sola reazione chimica.
Molto importante è anche il ruolo dell’acido linolenico (C18:3), la cui importanza e correlazione con il buon andamento della fertilità di una mandria è ben nota, ma che può influenzare negativamente il titolo di grasso.
Nel processo di saturazione di linoleico e linolenico si possono però formare degli intermedi che tendono a deprimere il tasso di grasso nel latte; un tipo di acido linoleico coniugato (CLA) molto pericoloso è il trans10cis12 linoleico, che viene anch’esso progressivamente saturato ma di cui bastano percentuali bassissime per inibire la sintesi endomammaria di grasso.
Figura 2 – Vie semplificate di bioidrogenazione dell’acido lineloico, che sottolineano la via principale t11 che coinvolge intermedi compatibili con il grasso del latte e la via minore t10 che coinvolge intermedi noti per inibire la sintesi di grasso del latte da parte della ghiandola mammaria. Adattato da Ferlay et al. (2017).

Un altro dei problemi causati dagli acidi grassi insaturi è la loro attività antibatterica, diversa a seconda della sensibilità delle specie batteriche e che si svolge a livello di disgregazione della membrana cellulare. Questa attività è molto più dannosa in caso di pH ruminale basso.
Detto questo, e dopo un’attenta analisi del rapporto c18:1/c18:0per verificare l’efficienza della via metabolica, la presenza di acido vaccenico ed il suo rapporto con i CLA, si può provare a diminuire l’apporto di acidi grassi insaturi della razione, soprattutto l’apporto di c18:2, considerando che gli UFA (unsaturated fatty acids) non dovrebbero mai superare il 50% del totale del grasso apportato.
Insomma, il problema titolo di grasso è assai complesso e non deve essere semplificato ma studiato con molta attenzione.





Scrivi un commento
Devi accedere, per commentare.